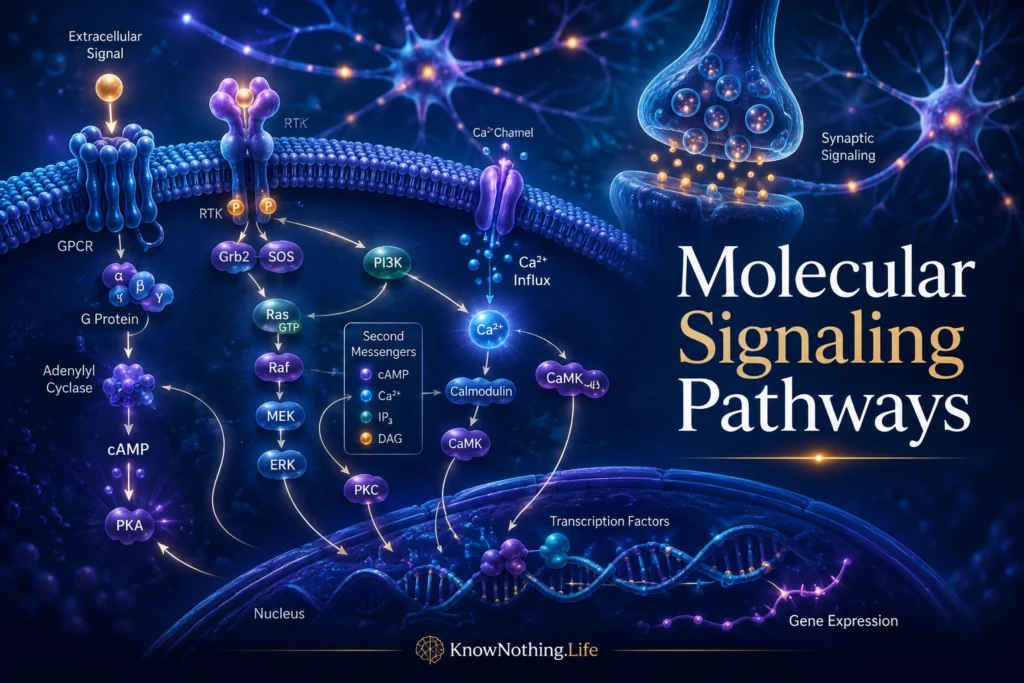
Molecular signaling pathways are organized chains of biochemical events that allow cells to receive information, process it, and respond. A signal may begin outside the cell, such as a hormone, neurotransmitter, growth factor, immune molecule, or sensory cue. It may also begin inside the cell, such as a change in calcium level, energy state, DNA damage, oxidative stress, or neural activity. Once detected, the signal is transmitted through proteins, enzymes, second messengers, receptors, kinases, phosphatases, transcription factors, and other molecular switches. In simple terms, a signaling pathway is the cell’s way of turning information into action.
These pathways are essential in every part of biology, but they are especially important in the nervous system. Neurons and glial cells use signaling pathways to regulate ion channels, synaptic strength, neurotransmitter release, gene expression, growth, survival, inflammation, myelination, metabolism, and memory-related plasticity. A neuron firing an action potential is not only producing an electrical event; it may also activate intracellular cascades that alter receptors, change protein synthesis, or influence transcription in the nucleus. NCBI’s overview of second messengers notes that neurons use many intracellular signals, each produced, removed, and targeted through different mechanisms.
Receptors: Where Signals Begin
Most molecular signaling pathways begin with receptors. Receptors are proteins that detect specific signals and convert them into cellular responses. Some receptors sit on the cell surface and bind molecules outside the cell. Others are located inside the cell and respond to molecules that can cross the membrane, such as steroid hormones. In the nervous system, receptors detect neurotransmitters, neuromodulators, neurotrophic factors, hormones, cytokines, and extracellular matrix signals. Their job is not merely to “receive” a message, but to translate that message into a biochemical change.
Receptors come in several major forms. Ionotropic receptors are ligand-gated ion channels that open quickly when a neurotransmitter binds, producing rapid electrical effects. G protein-coupled receptors, or GPCRs, activate intracellular G proteins and second messengers. Receptor tyrosine kinases, such as TrkB, can trigger phosphorylation cascades involved in growth, survival, and plasticity. Nuclear receptors can alter gene expression directly. The importance of GPCRs is especially broad; StatPearls describes them as activating intracellular signaling pathways, including generation of second messengers such as cyclic AMP and inositol phospholipids.
Second Messengers and Amplification
Second messengers are small intracellular molecules that carry and amplify signals inside cells. They allow an external signal to produce a larger internal response. Major second messengers include cyclic AMP, calcium ions, inositol trisphosphate, diacylglycerol, cyclic GMP, and nitric oxide. A neurotransmitter or hormone may bind to one receptor, but that receptor can activate enzymes that generate many second messenger molecules. This amplification allows small signals to create meaningful cellular changes.
The discovery of cyclic AMP was one of the foundations of modern signal transduction. Earl Sutherland showed that cyclic AMP could act as a “second messenger,” translating hormonal signals into intracellular effects. The Nobel Prize press release for Sutherland’s 1971 award described cyclic AMP as a second messenger in relation to hormonal effects, a discovery that opened wide biological perspectives. In neurons, second messengers are crucial because they connect synaptic activity to longer-lasting cellular responses. A brief neurotransmitter signal can produce changes in phosphorylation, receptor trafficking, gene expression, or synaptic strength that outlast the original signal.
G Proteins and Signal Transduction
G proteins are molecular switches that help transmit signals from receptors to intracellular targets. When a GPCR is activated, it can change the state of a G protein, allowing that G protein to influence enzymes or ion channels. Some G proteins stimulate adenylyl cyclase and increase cyclic AMP. Others inhibit cyclic AMP production or activate phospholipase C, leading to inositol trisphosphate, diacylglycerol, calcium release, and protein kinase C activation. This allows different receptors to produce different cellular effects even when they belong to the same broad receptor family.
Martin Rodbell and Alfred Gilman were awarded the 1994 Nobel Prize in Physiology or Medicine for their discovery of G proteins and their role in signal transduction. The Nobel Prize summary states that they discovered G proteins and the role of these proteins in cellular signal transduction. Their work mattered because it revealed that communication between a receptor and the cell interior often requires intermediate molecular switches. The signal does not simply enter the cell directly. It is relayed, modified, amplified, and controlled.
Protein Phosphorylation and Molecular Switches
One of the most important mechanisms in molecular signaling is phosphorylation, the addition of a phosphate group to a protein. Protein kinases add phosphate groups, while phosphatases remove them. This reversible process can change a protein’s shape, activity, location, stability, or interaction with other molecules. Reversible phosphorylation is one of biology’s most common regulatory mechanisms because it allows cells to turn processes on, turn them off, or tune them in graded ways.
Edmond Fischer and Edwin Krebs helped establish the central role of reversible protein phosphorylation. The Nobel Prize press release for their 1992 award notes that they characterized the first protein revealing a new mechanism for enzyme control through reversible phosphorylation. In the brain, phosphorylation affects ion channels, receptors, synaptic scaffolding proteins, transcription factors, enzymes, and cytoskeletal elements. A kinase cascade can therefore alter both immediate function and long-term cellular state. This is why molecular signaling pathways are often described as networks of switches rather than straight lines.
Kinase Cascades and MAPK Signaling
Kinase cascades are signaling pathways in which one kinase activates another, creating a chain of phosphorylation events. One of the best-known examples is the MAPK/ERK pathway, which can connect signals at the cell surface to changes in the nucleus. Growth factors, neurotrophins, and synaptic activity can activate Ras, Raf, MEK, ERK, and downstream transcriptional regulators. In neurons, MAPK signaling is often discussed in relation to development, synaptic plasticity, survival, and memory-related gene expression.
The power of a cascade is that it can amplify, filter, and shape signals. It can also create specificity. The same general pathway may produce different effects depending on cell type, timing, location, strength, scaffold proteins, and crosstalk with other pathways. This matters in neurons because a calcium signal in one dendritic spine may need to affect a local synapse, while a stronger or repeated signal may need to reach the nucleus and alter gene expression. Molecular signaling is therefore not just chemistry happening everywhere at once. It is chemistry organized in space and time.
Calcium Signaling in Neurons
Calcium is one of the most important signaling molecules in the nervous system. It enters neurons through voltage-gated calcium channels, NMDA receptors, and other channels, and it can also be released from intracellular stores. Calcium controls neurotransmitter release, synaptic plasticity, gene transcription, enzyme activity, mitochondrial function, and cell survival. Because calcium can be powerful and potentially dangerous, cells regulate it tightly through pumps, buffers, organelles, and localized signaling domains.
Calcium signaling is especially important at synapses. A brief calcium rise in a presynaptic terminal can trigger neurotransmitter release. In dendrites, calcium entry can help determine whether a synapse undergoes long-term potentiation or long-term depression. In the nucleus, calcium-dependent pathways can activate transcription factors such as CREB. This makes calcium a bridge between electrical activity and molecular change. When neurons fire, calcium can help translate that activity into plasticity, protein synthesis, and long-term adaptation.
CREB, Gene Expression, and Memory
Some signaling pathways end in the nucleus by changing gene expression. One of the most studied transcription factors in neuroscience is CREB, or cAMP response element-binding protein. CREB can be activated by pathways involving cyclic AMP, protein kinase A, calcium/calmodulin-dependent kinases, MAPK, and other signaling systems. Once activated, CREB helps regulate genes involved in synaptic growth, neuronal survival, plasticity, and long-term memory. A review by Sakamoto and colleagues describes CREB as a regulator that couples synaptic activity to long-term changes in neuronal plasticity, a process thought to underlie learning and memory.
Eric Kandel’s work helped connect molecular signaling pathways with memory storage. His review “The Molecular Biology of Memory Storage” described memory as a dialogue between genes and synapses, emphasizing that long-term memory depends on molecular changes that alter synaptic function. This is one of neuroscience’s central insights: learning is not only a psychological process. It involves biochemical pathways that convert experience into lasting cellular change.
Neurotrophins and BDNF-TrkB Signaling
Neurotrophins are growth-related signaling molecules that support neuronal survival, development, synaptic maturation, and plasticity. Brain-derived neurotrophic factor, or BDNF, is especially important in the nervous system. BDNF binds to the TrkB receptor, a receptor tyrosine kinase, and activates intracellular pathways including MAPK/ERK, PI3K-Akt, and PLCγ. These pathways can affect synaptic strength, protein synthesis, dendritic structure, survival, and gene expression.
BDNF-TrkB signaling is deeply connected to synaptic development and plasticity. A review by Yoshii and Constantine-Paton notes that BDNF-TrkB signaling is involved in transcription, translation, and protein trafficking during synaptic development. Later reviews emphasize that the timing and location of BDNF signaling matter, because the same molecule can have different effects depending on where and when it acts. This is a recurring theme in molecular signaling: context determines meaning. The pathway is not just what molecules are present, but where they are, when they activate, and what other signals are active at the same time.
Crosstalk, Feedback, and Network Logic
Molecular signaling pathways are often drawn as simple arrows, but real signaling is networked. Pathways interact with one another through crosstalk, feedback loops, feedforward loops, scaffolding proteins, compartmentalization, and local degradation. A signal that begins through a GPCR may influence kinase cascades, calcium signaling, transcription factors, and ion-channel function. A growth-factor pathway may interact with metabolic pathways, inflammatory pathways, and synaptic activity. Cells do not run one pathway at a time. They integrate many signals into one response.
Feedback is especially important. Negative feedback can shut down a response and prevent overactivation. Positive feedback can strengthen and stabilize a cellular state. In memory research, scientists have proposed that persistent molecular feedback loops may help maintain synaptic changes over time. A review on molecular mechanisms of the memory trace notes that long-term memory consolidation depends on synaptic plasticity and key molecular signaling cascades that strengthen particular synaptic connections. This network logic helps explain how brief experiences can sometimes produce lasting biological change.
Clinical Importance of Signaling Pathways
Many diseases involve altered molecular signaling pathways. Cancer often involves abnormal growth-factor signaling, kinase activation, or cell-cycle regulation. Neurodegenerative disorders may involve disrupted protein handling, inflammation, mitochondrial signaling, calcium regulation, or synaptic survival pathways. Psychiatric and neurodevelopmental conditions can involve altered receptor function, synaptic plasticity, gene regulation, immune signaling, or intracellular cascades. Signaling pathways also matter in addiction, chronic pain, epilepsy, depression, anxiety, traumatic brain injury, and stroke.
Many treatments work by targeting signaling. Some drugs affect receptors. Others affect enzymes, kinases, phosphodiesterases, ion channels, transporters, or inflammatory mediators. The growing importance of molecularly targeted therapies reflects a basic truth: disease often occurs not because a cell lacks information, but because it interprets or transmits information incorrectly. Understanding signaling pathways helps medicine become more precise, moving from broad symptom control toward specific molecular mechanisms.
Why Molecular Signaling Pathways Matter
Molecular signaling pathways matter because they explain how cells make decisions. A cell must know when to grow, divide, repair, fire, release neurotransmitter, strengthen a synapse, express a gene, conserve energy, respond to stress, or die. It does not make these decisions consciously. It uses molecular systems that detect signals, process them, and trigger responses. Signaling pathways are the biochemical logic of cellular life.
In the brain, these pathways become especially meaningful because they connect experience to biology. A sound, memory, drug, stressor, reward, injury, or lesson can alter neural activity, which can activate molecular cascades, which can change synapses, proteins, genes, and circuits. To understand molecular signaling pathways is to understand one of neuroscience’s deepest principles: the mind depends not only on neurons firing, but on cells translating signals into lasting biological change.